
Tracing Ancient Shorelines: Field Notes from Java on Early Human Evolution
In mid-February, I returned to Southeast Asia—this time to Indonesia—together with Professor Erika Schagatay. At the start of our trip, we met up with Professor José Joordens, a Dutch anthropologist and paleoanthropologist. With her, we visited several excavation sites where she has been working, as well as museums in central Java.
José Joordens became widely known for her work on the engraved freshwater Pseudodon shell from Trinil—a remarkable artifact originally collected by Eugène Dubois and his team in the 1890s and later kept in Dutch museums.
The zig-zag engraving on one of these shells went unnoticed for more than a century. It was in fact paleoanthropologist Stephen Munro who recognized the engraving already in 2007, while examining photographs he had taken of the shell collection in Naturalis Biodiversity Center. The finding was later formally described in 2014. The carving has been dated to between approximately 430,000 and 540,000 years ago, making it the oldest known example of an abstract or symbolic marking created by any human.
Along the Solo River and the World of Homo erectus
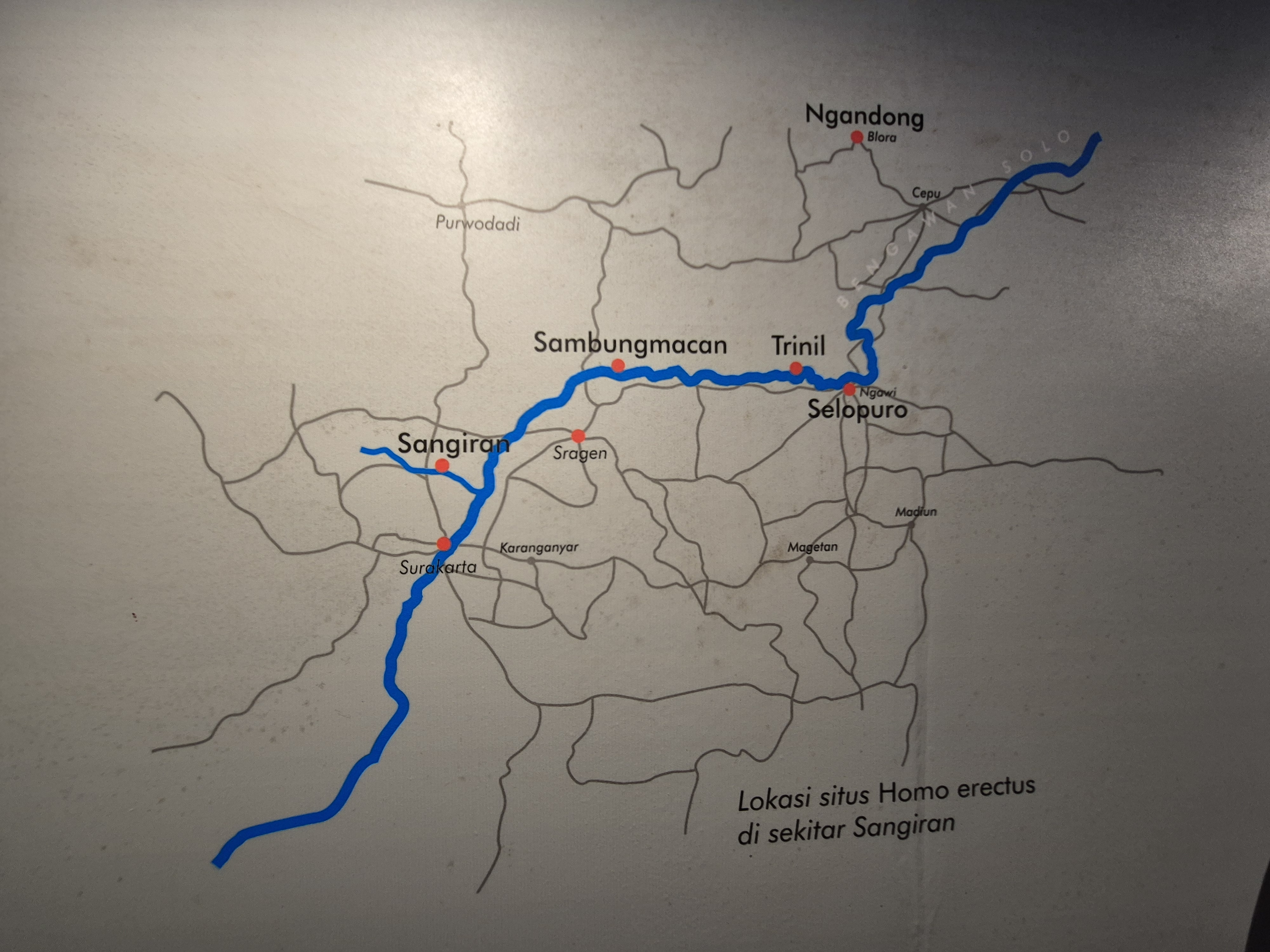
The Solo River basin is one of the richest regions in the world for Homo erectus discoveries. Groups of H. erectus lived along this river system from roughly 2 million years ago until as recently as 100,000 years ago, including the well-known Ngandong population—among the latest surviving Homo erectus known.
During our trip, Erika and José also met with numerous local researchers as part of early planning for a future interdisciplinary project on the Bajau Laut (Sama-Bajau) communities. The project aims to explore diving adaptations from physiological, anthropological, and cultural perspectives.
José has a long-standing interest in the waterside hypothesis and suggests that Homo erectus may have been shallow-water divers and foragers. The argument is supported by evidence such as thick cortical bones, a relatively dense skeleton, breath-hold capacity, and repeated associations with shell-bearing or riverine sites. The Trinil shell assemblage is central here, and it has also shown that shells were used as tools. In addition, José has excavated in Kenya’s Turkana Basin—another key region for early hominin interactions with aquatic environments.
Visiting Trinil: Dubois’s Historic Site
One of the highlights of our journey was visiting Trinil, where Eugène Dubois uncovered the first Homo erectus fossils—his famous “Java Man.” It was also here that the engraved shell was found. Local people still consider the site haunted, and many avoid being there at night. During Dubois’s excavations, prisoners were used as laborers, and a number of them died during the work, reportedly chained together and forced to excavate under horrific conditions.
We met long-time collaborators of José who have worked at Trinil for years. We also had access to a back room where numerous remnants, casts, and planning materials are kept, including a copy of the engraved Pseudodon shell. In the surrounding area, we also explored exhibitions and museums that highlight the life and environment of Homo erectus. Standing by the Solo River—where the earliest discoveries of Java Man were made, alongside men fishing with large handheld nets—brought a quiet sense of continuity between past and present.











A Waterside Landscape Two Million Years Ago
Java was a very different place two million years ago. The region was shaped by extensive river systems, lakes, wetlands, and coastal environments, and supported a wide range of mammals adapted to aquatic or semi-aquatic niches—such as hippos, elephants, and other water-associated species. Humans were part of this landscape, and this is also the region where Homo erectus appears to have persisted the longest.
Fossils attributed to Homo erectus appear in Africa at around two million years ago, including recent finds from sites such as Drimolen in South Africa, while the well-known Dmanisi material in Georgia dates to around 1.8 million years ago. In Southeast Asia, the Java record begins around 1.8 million years ago and continues until approximately 100,000 years ago.
Over this immense timespan, H. erectus brains increased in size—a gradual but significant development that may reflect changing environments and a long-term reliance on nutrient-rich aquatic and waterside resources.
In other parts of the world, Homo erectus either evolved into other lineages (for example through Homo heidelbergensis, often discussed as a common ancestor of Homo sapiens, Neanderthals, and Denisovans) or went extinct. Yet on isolated islands in Southeast Asia—such as Flores, Luzon, and Sulawesi—human species appear to have persisted much longer.
Water, Diving, and Human Evolution
Homo erectus had characteristically thick bones and may have been skilled divers. According to some researchers—especially Marc Verhaegen—the species’ history of breath-hold diving shaped both biology and behavior. Verhaegen argues that:
- Homo erectus frequently exploited coastal, riverine, and lake environments.
- Breath-hold diving may have been used to collect shellfish, aquatic plants, and other high-value foods.
- Their relatively heavy and dense skeleton may have functioned as natural ballast during shallow diving.
- In addition to diving, Homo erectus may often have floated on the back while foraging at the surface—somewhat comparable to modern sea otters—allowing extended time in the water with minimal energy expenditure.
- Long-term reliance on aquatic resources could have influenced brain growth and broader physiology.
- Cranial features such as paranasal air sinuses have been discussed in relation to pressure regulation and repeated submersion, though interpretations remain debated.
- Other traits sometimes highlighted include the ability to suck shellfish from shells and hypersensitive fingertips, well suited for detecting hidden prey while foraging underwater.
Homo sapiens later continued to exploit marine foods, which is significant given our need for DHA fatty acids to support a large brain. Although our species eventually became less dependent on diving, technological development likely allowed us to harvest aquatic resources in new and more efficient ways. Coastal foraging probably remained crucial even as our bodies became lighter and more specialized for long-distance walking and endurance running.
Average brain size in the human lineage increased steadily until roughly 300,000–200,000 years ago, after which it appears to have plateaued. A gradual reduction in average brain size becomes visible later, beginning around 50,000 years ago and continuing into more recent prehistory.
Given the high energetic demands of large brains, it seems likely that nutrient-rich foods centered on marine and aquatic resources played an important role during the period of brain expansion. The later reduction in brain size may reflect a gradual decrease in reliance on these resources as humans increasingly diversified their diets and food sources. Many large-brained mammals are closely tied to aquatic food chains during their evolution, which helps explain why the relationship between water, diet, and brain evolution remains such an intriguing topic.
Interestingly, some of the museums we visited—such as the Sangiran Museum Site—explicitly embraced the idea that Homo erectus lived close to water and may represent the earliest island-adapted human. Seeing this interpretation presented so openly was refreshing.
Engraved Shells and Shared Human Cognition
The Trinil shell itself is fascinating. Its zig-zag pattern resembles markings found in other early human contexts, such as the cross-hatched engravings made by early Homo sapiens on pieces of red ochre at Blombos Cave in South Africa (dated to about 73,000 years ago), as well as early Neanderthal engravings, including the cross-hatched markings from Gorham’s Cave in Gibraltar (~39,000 years old).
These similarities raise intriguing possibilities about shared cognitive tendencies—suggesting that geometric pattern-making may reflect deep underlying structures of human and pre-human thought. It also points to a long period when abstract marks existed without figurative art, before a later shift within Homo sapiens—after our species had already been around for more than 200,000 years.
Final Reflections
Traveling with José was inspiring. She is one of the few paleoanthropologists who treats the waterside hypothesis with genuine scientific curiosity, and her background as a marine biologist adds depth to her perspectives.
It is also striking that the earliest known sign of abstract behavior—an intentional engraving—comes not from stone tools or cave walls, but from a freshwater shell, further highlighting the importance of waterside environments in our evolutionary history.



It is so great to hear how much work is being done in and on this fascinating area Erik. Our connection with the Marginal World Aquatic clearly stretches back and informs our deep evolutionary development much more than widely understood.
February 20, 2026 at 8:04 am
Hi Andrea,
Thank you for your comment and for engaging with the post. Yes, a great deal of work is being done in this area, and it is becoming clearer than ever that we have a deep evolutionary connection to aquatic and waterside environments. Unfortunately, this connection is still largely unknown.
The mainstream narrative continues to emphasize a savanna-based story, where humans evolved primarily through cognitive problem-solving in harsh terrestrial environments. Yet much of what has shaped us—our breathing, bodily movements, sensory systems, and even aspects of our thinking—likely developed in close interaction with water-rich landscapes and a more embodied sense of being part of the natural world.
Best regards,
Erik
February 20, 2026 at 8:44 am
Many many thanks for this beautiful website, Erik! 🙂
Fossils attr.to H.erectus appear in Africa c.2 Ma, yes, but is this attribution correct? Endo-retroviral evidence (humans & Asian mammals lack PtERV1 DNA) suggests our ancestors were not in Africa then? Most likely, African australopiths were closer relatives of Gorilla or Pan than of Homo?
February 20, 2026 at 1:40 pm
Hi Marc,
Many thanks for your thoughtful comment — and for engaging with the post. 🙂
You raise a very important and still unresolved issue. While fossils attributed to Homo erectus do appear in Africa around ~2 million years ago, the question of whether these remains are correctly classified is far from settled. As you point out, the endo-retroviral evidence — particularly the absence of PtERV1 sequences in humans and Asian mammals, contrasted with their presence in African apes — complicates a straightforward African origin narrative for our lineage at that time.
I agree that this opens up the possibility that several African australopiths may represent closer relatives of Gorilla or Pan rather than direct ancestors of Homo. At the very least, it suggests a more branching and reticulate evolutionary history than is often presented, with multiple hominin lineages adapting to different ecological niches.
What remains particularly striking to me is that, regardless of where Homo erectus first emerged, it is in Southeast Asia that we see its longest persistence, clear ecological success, and substantial brain expansion. This strongly suggests that aquatic and waterside adaptations were central to its way of life — and that Asia played a much more significant role in human evolution than is usually acknowledged. In this light, I find the hominin record from Flores, Luzon, and Sulawesi especially intriguing. These populations suggest that multiple hominin lineages were established in Asia early on, possibly derived from H. erectus ancestors or closely related sister groups. Taken together, this points toward a more complex and persistent Asian hominin presence than a straightforward “Out of Africa” dispersal model would imply.
I very much appreciate you bringing the retroviral evidence into the discussion — it’s exactly this kind of cross-disciplinary perspective we need.
Warm regards,
Erik
February 23, 2026 at 9:44 am
FYI, our recent ideas:
–Evolution of Bipedality, google “aquarboreal”
–African Ape evolution, google “Vaneechoutte Australopithecus”
–Language evolution, google “Seafood, diving, song and speech”
–Hominoid evolution book, google “GondwanaTalks Verhaegen”
–Archaic Homo evolution, google “Homo shellfish diving”
March 12, 2026 at 11:44 pm
Hi Erik, thanks for the interesting website and information.
I would like to share something I found out recently. Maybe some of you knew already, but I have never read it before. It concerns the paradox of the small-brained Sirenians (dugongs and manatees). The fact is that their diet, mainly sea grass, does not contain arachidonic acid (ArA) and docosahexaenoic acid (DHA). Mollusks are a rich source of these LC-PUFAs because they feed upon the marine unicellular eukaryotes which are the only producers (except some bacteria) of ArA and DHA.
It turns out that the small brains of the Sirenians, often used as an argument against our semi-aquatic past, confirm the need for ArA and DHA which are essentially marine in origin.
February 21, 2026 at 6:09 pm
Hi Mario,
Good to hear from you, and thank you for sharing this — it’s a new point to me and a very interesting and important one.
You’re absolutely right that sirenians are often brought up in discussions against a semi-aquatic phase in human evolution, precisely because of their relatively small brains despite a fully aquatic lifestyle. But as you point out, their case actually highlights something crucial: diet matters as much as habitat.
Sirenians feed primarily on seagrass, which is poor in DHA and ArA. In contrast, for hominins exploiting shellfish, fish, and other aquatic fauna, access to these long-chain fatty acids would have been consistent and reliable.
So rather than undermining the idea of a semi-aquatic or waterside past, the sirenian example actually refines it — reminding us that not all aquatic niches exert the same evolutionary pressures.
Warm regards,
Erik
February 23, 2026 at 9:56 am
Thank you for your additional comments, Erik. In fact, you bring up something most important about diet: consistency/continuity. The growing fetal and infant brain is not only high in demands for energy (glucose, and ketones from fatty acids) and for specific brain cell membrane building blocks (long chain polyunsaturated fatty acids such as ArA and DHA), but the most important factor to enable such disproportionally extreme increase in brain volume might be: uninterrupted supply or continuity. The only habitat/econiche where the three conditions of 1) easily available energy (fatty acids), 2) large amounts of brain specific nutrients (LC-PUFAs, taurine, choline, magnesium, selenium, iodine) and 3) continuous, uninterrupted supply of energy and building blocks are met, are, of course, coastal/riparian zones. Animal brains from predation are rich in DHA, but their supply is uncertain (seasonal = discontinued) and difficult to access, because the prey is dispersed (and … runs away) and because of competition with fierce predators (giant hyenas and saber tooth tigers during the Pleistocene).
The extremely large brains of large mammals, such as those of pinnipeds, cetaceans and humans, are only achievable when the supply of energy and building blocks is constant. The only environment where such brains could have evolved is marine/riparian/lacustrine.
Or, as ChatGPT would put it (after some insistance):
Logical schema (premise → conclusion)
Premise 1 (Demand): Human infant/child brain development has an exceptionally high and prolonged metabolic cost, implying that brain growth is constrained by sustained substrate delivery (not just occasional “peaks” of intake). (Kuzawa et al. 2014).
Premise 2 (Dual constraint): Normal human brain development faces two coupled constraints: (i) very high energy requirement, and (ii) reliable access to brain-selective nutrients needed for structure and function (e.g., DHA, ArA, iodine, iron, zinc, cupper, selenium, taurine) (Cunnane & Crawford 2014).
Premise 3 (Buffering mechanism): Hominins partly solve the energy side via maternal/foetal fat buffering (third-trimester foetal fat + postnatal fat) that supports ketone availability and provides a reserve of DHA—i.e., buffering is explicitly framed as a continuity solution (Cunnane et al. 1993; Cunnane & Crawford 2003, 2014).
Premise 4 (Milk is not the main differentiator): Great-ape milk and human milk are broadly similar in macronutrient patterning; therefore, a large human-specific divergence in infant brain growth is unlikely to be explained primarily by “milk recipe” alone and instead points to differences in overall continuity of maternal nutrient throughput and buffering capacity across pregnancy + lactation + early childhood (Garcia et al. 2017). The DHA content of human milk depends on the acute fish intake (Lauritzen et al. 1999).
Premise 5 (Continuity depends on maternal diet ecology): Key brain lipids in milk—especially DHA—vary widely between populations and are strongly associated with maternal access to DHA-rich foods; the highest milk DHA values are reported mainly in coastal populations and are associated with marine food consumption (Innis et al. 2007,2014). Therefore, ecology that enables frequent DHA access improves continuity of brain-building inputs via maternal tissues and milk (Brenna et al. 2007).
Premise 6 (Why water-bound foods are a strong continuity candidate): Shore/river/lake food webs provide dense packages of brain-selective nutrients; and shore foraging can yield very high caloric return rates (even if tide-limited), consistent with a niche that can lower variance in maternal/juvenile energy intake when combined with complementary foods (Cunnane & Crawford 2014; De Vynck et al. 2016).
Conclusion (Most plausible ecological inference under these premises): If (i) large human infant brain expansion requires continuous energy plus brain-building substrates, (ii) milk macronutrients are not uniquely different across hominoids, and (iii) maternal DHA (and associated micronutrient) availability tracks ecology—with the richest known “brain-selective nutrient” packages concentrated in aquatic food webs—then the most plausible ecological setting enabling the required maternal buffering/continuity is a water-bound foraging niche (coastal/estuarine/freshwater), rather than a purely terrestrial, high-variance food base (Cunnane & Crawford 2014; Brenna et al. 2007).
References
Brenna JT, Varamini B, Jensen RG, et al. Docosahexaenoic and arachidonic acid concentrations in human breast milk worldwide. Am J Clin Nutr 2007;85(6):1457–1464. https://doi.org/10.1093/ajcn/85.6.1457.
Cunnane SC, et al. The importance of energy and nutrient supply in human brain evolution. Nutr Health 1993. (PubMed: https://pubmed.ncbi.nlm.nih.gov/8183489/)
Cunnane SC, Crawford MA. Energetic and nutritional constraints on infant brain development: Implications for brain expansion during human evolution. J Hum Evol 2014;77:88–98. https://doi.org/10.1016/j.jhevol.2014.05.001.
Cunnane SC, Crawford MA. Survival of the fattest: fat babies were the key to evolution of the large human brain. Comp Biochem Physiol A Mol Integr Physiol 2003;136:17–26. https://doi.org/10.1016/S1095-6433(03)00048-5.
De Vynck JC, Anderson R, Atwater C, et al. Return rates from intertidal foraging from Blombos Cave to Pinnacle Point: Understanding early human economies. J Hum Evol 2016;92:101–115. https://doi.org/10.1016/j.jhevol.2016.01.008.
Garcia M, Power ML, Moyes KM. Immunoglobulin A and nutrients in milk from great apes throughout lactation. Am J Primatol 2017;79(3):1–11. https://doi.org/10.1002/ajp.22614.
Innis SM. Human milk: maternal dietary lipids and infant development. Proc Nutr Soc 2007. (PubMed: https://pubmed.ncbi.nlm.nih.gov/17637092/)
Innis SM. Impact of maternal diet on human milk composition and neurological development of infants. Am J Clin Nutr 2014;99:734S–741S. https://doi.org/10.3945/ajcn.113.072595.
Kuzawa CW, Chugani HT, Grossman LI, et al. Metabolic costs and evolutionary implications of human brain development. Proc Natl Acad Sci U S A 2014;111(36):13010–13015. https://doi.org/10.1073/pnas.1323099111.
Lauritzen L, Jøgensen MH, Michaelsen KF. The docosahexaenoic acid (DHA) content of human milk depends on the acute fish intake. Pediatr Res 1999;45:916. https://doi.org/10.1203/00006450-199906000-00191.
February 26, 2026 at 10:15 am
Hi Mario,
Thanks for your comment. Yes, consistency is key here. Humans must have continued to rely on marine and aquatic resources for a long time. Since Homo sapiens developed even larger brains than Homo erectus, it seems clear that these resources remained important, even if our species was no longer as dependent on diving itself. Instead, people may increasingly have exploited shorelines by walking long distances along coasts (combined with swimming and some level of diving, of course, as suggested by for example the prevalence of surfer’s ear in Homo sapiens) and by utilizing aquatic resources differently and more efficiently thanks to new technologies.
This idea of continuity is also reflected in the material culture of early Homo sapiens and our ancestors, which remained remarkably persistent over long periods of time—for example in Middle Stone Age tool traditions. People appear to have lived in broadly similar ways for many millennia. A relatively stable lifestyle combined with increasing brain size suggests to me that the function of the expanding brain may not have been primarily creativity, intelligence, or proactive communication, as is often assumed in the current paradigm.
Rather, it may have supported stability and the reliable reproduction of behaviors and strategies—such as tool use, landscape navigation, and the transmission of practical knowledge across generations. Tools themselves can be seen as an exosomatic extension of the body, with the growing brain gradually incorporating them into everyday behavior. Similarly, navigating large landscapes in search of food resources and suitable tool-making materials — such as particular types of stone — required memory, orientation, and coordination. In my view, singing played a role in enabling this kind of long-distance movement, the repetition of coordinated hand movements, the incorporation of tools as part of the body, and social coordination.
In that sense, I am interested in a broader reinterpretation of the role of the evolving human brain. We often emphasize intellectual creativity, but the archaeological record—remarkably stable even within our own species—suggests that the brain may have been equally important for maintaining continuity, embodied knowledge, and long-term strategies for living within particular environments. In marine mammals as well, increasing brain size appears to be linked less to creativity and more to social bonding, synchronization, navigation, and vocal communication such as singing. There is therefore a need to rethink the role of our large brain in human evolution.
What do you think, Mario? You have been at the forefront of emphasizing the role of singing, as well as imitation and the importance of mirror neurons in our evolution.
Best regards,
Erik
March 8, 2026 at 3:32 pm
Well, lots of interesting inputs, Erik.
First, I agree that long term continuity (over generations) was essential to understand human evolution. But I especially wanted to emphasize the daily continuity of supply of energy (fatty acids, ketones) and building blocks (omega3 fatty acids), something only possible at the coast: rather safe with regard to predators, plenty of food, plenty of easy food, plenty of brain nutritional food (poly-unsaturated long chain fatty acids (omega3)), but most important: continuous supply. No periods of starvation, which are deleterious for brain development, but which are experienced by most terrestrial animals as they depend on an occasional successful hunt or on seasonally available fruits (e.g., chimps) or on any other uncertain source of nutrition. The only econiche that enabled the (preposterous!?) brain growth as we see it in humans and marine mammals is the sea and the coast, because of richness in brain food, but most of all because of certified continuity of supply.
That we were not in need for such preposterously large brains is clear when you consider that most animals (including our closest relatives) can do well with much smaller brains, and when you consider that our lineage has been on the brink of extinction several times: our large brain was not such an asset as we think it is: our species went almost extinct a few times only a million years ago, when we already had such large brains. And after all, despite our large brains, half of the world population is ruled by people like …, several of them even elected democratically.
Singing. Indeed. In fact, it is intriguing to find out why the human has such a large brain, why it is naked, why it can hold breath, why it is walking on hind legs and upright (only penguins can do that as well, waggling), and why it has so many other weird unprimate-like characteristics, but the essential question is: why can the human being speak!? All the other aspects are most weird indeed, until you realize what Alister Hardy and Elaine Morgan learned us. But in the end, speech is what eventually caused us to be over 8 billion, destroying our planet.
An animal that can tell another animal about the world, is at an advantage, of course. But if the explanation for our speech goes like: ‘we developed speech because it was advantageous’, you should ask why only our species developed speech, instead of every animal. That was my question, before I had ever heard about our possible aquatic past. (It was Marc Verhaegen who contacted me after having read my joint publication with John Skoyles about song as the predecessor for speech, an idea that I thought was very original, but that had been formulated already (although vaguely) by Darwin and since then by several others (more explicitly) before me. I gave up the self-righteousness that my original insights are original.
So, why can we speak? My conviction is that we can speak because we can sing. That brings us to the next question: Why can we sing? My conviction is that we can sing (and dance, unseparably connected to song) because we have an evolutionary – semi-aquatic – past of moving freely (freely is important here!) in three dimensions. Not as in trees (primates) – which is not freely, but as in air (song birds and bats) and as in water (cetaceans, pinnipeds). The fact that song is present only in highly mobile species was already noted by Janik & Slater (1997), although their interpretation of this observation differs from mine. I think that song becomes possible because of the ability of freely moving animals to mimick others. These animals must be able to mimick the movements of other animals IMMEDIATELY. In fact, it means that they must be able to predict the immediate future, based on what happened the moment before. And they are indeed able to do so (Just watch a flock of starlings or a school of fish).
Song is about mimicking overheard sounds. Gibbons are excellent ‘singers’ but they do not learn song by mimicking, their ‘song’ is innate. It is a complex call, not real song (Janik & Slater, 1997).
So, why can we sing? Because of our aquatic past (of course): it gave us control of breath (not possible for other primates) and it gave us mimicking, something animals in water (fish, pinnipeds, cetaceans) can do well to synchronize their movements, to escape predators, to chase prey, all in three dimensions. Mimicking is equal to predicting: calculating from what happened just before to predict what will happen the next milliseconds. Song and speech are about predicting what will come next. (There are a million songs in my head (slight exaggeration, apologies) of which I am able to predict each next note, and every rhythm change. I am sooo fantastic, and so are you.) I enjoy the way my grand children are dancing and singing from the age of one onwards. They simply can’t be stopped to move on rhythm. It is so natural, why do we overlook that aspect of our being?
It all fits (in my opinion of course). We can speak (always with gesture), because we can sing (and dance). And we can sing and dance, because we had an aquatic past (underwater swimming, deep diving, holding breath, predicting movement, mimicking). And of course, a large brain (only seen in aquatic mammals of our size) helps to develop language from song and is only possible because of plenty and continuous brain food, at the sea.
Vaneechoutte M. 2014. The origin of articulate language revisited: The potential of a semi-aquatic past of human ancestors to explain the origin of human musicality and articulate language. Human Evol. 29: 1-33. doi.
Vaneechoutte M, Munro S, Verhaegen M. Seafood, diving, song and speech. In: Vaneechoutte, M., A. Kuliukas, and M. Verhaegen. Was Man more aquatic in the Past? Fifty years after Alister Hardy. Waterside hypotheses of human evolution. Bentham Publishers. e-book; 2011; Chapter 12:181-189. https://users.ugent.be/~mvaneech/Vaneechoutte%20et%20al.%202011.%20Seafood,%20diving,%20song%20and%20speech.pdf
Vaneechoutte M, Skoyles JR. The memetic origin of language: modern humans as musical primates. J Memetics & Evol Models Inform Transm 1998;2.
https://users.ugent.be/~mvaneech/Vaneechoutte%20&%20Skoyles.%202000.%20Origin%20of%20language.pdf
Janik VMT, Slater PJB. Vocal learning in mammals. Adv Study Behav 1997;26:59-99, cited in:
Janik VM, Slater PJB. The different roles of social learning in vocal communication. Animal Behav 2000;60:1–11. https://pubmed.ncbi.nlm.nih.gov/10924198/
March 17, 2026 at 7:37 pm